
DOI:
https://doi.org/10.14483/udistrital.jour.colomb.for.2014.2.a06Publicado:
01-07-2014Número:
Vol. 17 Núm. 2 (2014): Julio-DiciembreSección:
Artículos de investigación científica y tecnológicaEstudio comparativo de la composición florística, estructura y diversidad de fustales en dos ecosistemas del campo de producción 50k cpo-09, llanos del orinoco colombiano
A comparative study of floristic composition, structure and pole diversity in two ecosystems at the 50k CPO-09 oil field in the Orinoco region of Colombia
Palabras clave:
biodiversity, forests, Colombia, conservation, ecology, endangered species, fragmentation (en).Palabras clave:
biodiversidad, bosques, Colombia, conservación, ecología, especies amenazadas, fragmentación (es).Descargas
Referencias
Andrade-C., M.G. (2011). Estado de conocimiento de la biodiversidad en Colombia y sus amenazas. Consideraciones para fortalecer la interacción ciencia-política. Rev. Acad. Colomb. Cienc., 35(137), 491-507.
Becerra, J. (1971). Notas de Ecología Forestal. Universidad Distrital Francisco José de Caldas. Bogotá D.C. Colombia.
Bernal, R., Celis, M. & S.R., Gradstein. (2007). Plant diversity of Colombia catalogued. Taxon 56 (1): 273.
Cárdenas, D., Giraldo-Cañas, G. y C. Arias. 1997. Vegetación. Capítulo 5. En: Instituto Geográfico Agustín Codazzi (IGAC). Zonificación Ambiental para el Plan Modelo Colombo-Brasilero (Eje Apaporis-Tabatinga: PAT). Bogotá, pp 183 - 228.
Cárdenas-Torres, M.A. (2011). Hábitat y distribución. En, M. Merchán (ed.): Historia natural y conservación del caimán llanero (Crocodylus intermedius) en Colombia (pp. 57-123). Madrid, España: Asociación Chelonia.
Carvajal, L., Patarroyo, J., Puentes, D. y J. Valero. (2007). Caracterización florística y estructural del piedemonte llanero en el departamento del Meta. Corporación para el Desarrollo Sostenible del Área de Manejo Especial La Macarena – Cormacarena, Universidad Distrital Francisco José de Caldas. Bogotá. 130 p.
Carvajal L. y J. Murillo. (2007). Análisis Florístico y Fitogeográfico del sector nororiental de la Sierra de la Macarena, Colombia. 214 p.
Duque A.J., D. Cárdenas y N. Rodríguez. (2003). Dominancia florística y variabilidad estructural en bosques de tierra firme en el noroccidente de la Amazonia Colombiana. Caldasia 25: 139-152.
Ferriol, M. y H. Marle. (2012). Los componentes alfa, beta y gamma de la biodiversidad. Aplicación al estudio de comunidades vegetales. Universidad Politécnica de Valencia. 10 p.
Holdridge, L.R. (1978). Ecología basada en zonas de vida. Instituto Interamericano de Cooperación para la Agricultura (IICA), San José, Costa Rica.
IBM Corporation. (2013). SPSS Statistics 19. U.S.A. IBM Corporation. ISBN: 3-900051-07-0, recuperado de http://www-01.ibm.com/software/co/analytics/spss/
Lamprecht, H. (1990). Silvicultura en los Trópicos. GTZ. República Federal Alemana. 64 - 92.
Maldonado-Ocampo J.A., Vari, R.P. y J.S. Usma. (2008). Checklist of the Freshwater Fishes of Colombia. Biota Colombiana 9 (2) 143 - 237, 2008.
Margalef. R. (1969). El ecosistema pelágico del Mar Caribe. Memoria Fund. La Salle Cien. Nat., 5-36.
Margalef, R. (1995). Ecología. Omega. Barcelona, España. 951 p.
Melo, O. y R. Vargas. (2003). Evaluación ecológica y silvicultural de ecosistemas boscosos,
Universidad del Tolima, CRQ, Carder, Corpocaldas, Cortolima, 238 p.
Ministerio de Ambiente, Vivienda y Desarrollo Territorial. (2010). Resolución 383 de 2010. Bogotá. 29 p.
Patil, G.P. & C. Taile. (1982). Diversity as a concept and its measurement, Journal of the American Statistical Association 77, 548-567.
Ramírez, C. y C. Kleinn. (2001). Inventario Forestal Global – GFS. Estudio Piloto en Costa Rica, Manual de Campo. Centro Agronómico Tropical de Investigación y Enseñanza (CATIE), Costa Rica, FAO, Ministerio del Ambiente y Energía (MINAE), Costa Rica. 36 p.
Rangel J.O. y A. Velásquez. (1997). Métodos de estudio de la vegetación. En, J.O. Rangel, P.D. Lowey y Aguilar, M. Colombia- Diversidad Biótica II. Bogotá.
Remsen, J.V., Jr., Cadena, C.D. Jaramillo, Nores, A.M. Pacheco, J.F. Pérez-Emán, J. Robbins, M.B. Stiles, F.G. Stotz, D.F. y K.J. Zimmer. (2012). A classification of the bird species of South America. American Ornithologists Union.
Sarmiento, J. (2007). La Familia Orchidaceae en Colombia. Actual Biol (Supl. 1): 84
Unesco. (1980). Ecosistemas de los Bosques Tropicales, Organización de las Naciones Unidas para la Educación, la Ciencia y la Cultura. París. pp 126-163.
Villareal H., Álvarez, M., Córdoba, S., Escobar, F., Fagua, G., Gast, H., Mendoza, F., Ospina, M. y A.M. Umaña. (2004). Manual de métodos para el desarrollo de inventarios de biodiversidad. Programa de Inventarios de Biodiversidad. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt. Bogotá, Colombia. 236 p.
Zar, J.H. (1984). Biostatistical analysis. Second edition. Prentice-Hall.
Cómo citar
APA
ACM
ACS
ABNT
Chicago
Harvard
IEEE
MLA
Turabian
Vancouver
Descargar cita
 |
Cárdenas - Torres M. A. (2014) Estudio comparativo de la composición florística, estructura y diversidad de fustales en dos ecosistemas del campo de producción 50k CPO-09, llanos del orinoco colombiano . Colombia Forestal, 17(2), 203-229. |
DOI: http://dx.doi.org/10.14483/udistrital.jour.colomb.for.2014.2.a06
ESTUDIO COMPARATIVO DE LA COMPOSICIÓN FLORÍSTICA, ESTRUCTURA Y DIVERSIDAD DE FUSTALES EN DOS ECOSISTEMAS DEL CAMPO DE PRODUCCIÓN 50k CPO-09, LLANOS DEL ORINOCO COLOMBIANO
A comparative study of floristic composition, structure and pole diversity in two ecosystems at the 50k CPO-09 oil field in the Orinoco region of Colombia
Miguel Andrés Cárdenas-Torres1
1 Consorcio Megaoil y Asociación Chelonia, Bogotá D.C., Colombia. miguelandrescardenas@gmail.com
Recepción: 12 de marzo de 2014 / Aprobación: 17 de junio de 2014
RESUMEN
En este trabajo se compara la estructura y composición de fustales en dos ecosistemas de la Orinoquia colombiana sobre un área de 10 465 hectáreas. Fue elaborado un mapa de coberturas a escala 1:10 000 para cartografiar el bosque de galería y el bosque denso alto de tierra firme, con el fin de implementar en campo un muestreo aleatorio estratificado, donde cada unidad muestral se seleccionó completamente al azar. Se recolectó información en 16 unidades de muestreo (parcelas) de 0.1 ha en ambos ecosistemas (8 en cada uno), incluyendo información de los individuos con diámetro a la altura del pecho (DAP) mayor a 10 cm y tomando datos de especie, circunferencia a la altura del pecho (CAP), altura total y a la base de la copa. Se encontró una mayor riqueza total de especies en bosque de galería (81) que en bosque denso alto de tierra firme (76); sin embargo, este último arroja una mayor heterogeneidad y una mejor estructura vertical. Entre ambos ecosistemas incluyen la presencia de 12 especies amenazadas en las diferentes categorías según las bases de datos de los Libros Rojos de Colombia, la Resolución 0192 de 2014 de Colombia, la RedList de la Unión Internacional para la Conservación de la Naturaleza (UICN) y la Convención sobre el Comercio Internacional de Especies Amenazadas de Fauna y Flora Silvestres (CITES).
Palabras clave: biodiversidad, bosques, Colombia, conservación, ecología, especies amenazadas, fragmentación.
ABSTRACT
A structure and composition study of poles in two wooded areas of Colombian Orinoquia totalling 10 465 hectares is presented. A 1: 10 000 scale map was made for mapping gallery forest and high dense forest from tierra firma, in order to implement a field stratified sampling, in which each sample unit is selected randomly. Data from 16 sampling units of 0.1 ha each from both ecosystems (for each of them) were collected, and included information about individuals with diameter at breast height(DBH) more than 10 cm and taking data of species, circumference at breast height (CBH), total height and height to crown base . Higher species richness was found in gallery forest with 81 species whereas tierra firma had 76. The latter however, had greater heterogeneity and a better vertical structure. Both ecosystems included the presence of 12 endangered species in different categories according to the Colombia red data book, Resolution 0192 of 2014 of Colombia, IUCN and CITES.
Key words: biodiversity, forests, Colombia, conservation, ecology, endangered species, fragmentation.
INTRODUCCIÓN
Colombia es el primer país del mundo en diversidad de aves y orquídeas (Remsen et al., 2012; Sarmiento, 2007), segundo en plantas, anfibios, peces dulce-acuícolas y mariposas (Bernal et al., 2007; Maldonado-Ocampo J.A et al., 2008; Andrade-C, 2011), tercero en diversidad de reptiles y palmas y cuarto en mamíferos (Andrade-C, 2011). Según datos oficiales disponibles en el Sistema de Información sobre Biodiversidad de Colombia, la región de la Orinoquia colombiana (con 300 000 km2) cuenta con un total de 92 especies de aves acuáticas, 108 anfibios, 119 reptiles, 318 mamíferos y 2692 plantas.
Esta región se encuentra conformada por enormes extensiones de sabanas naturales, áreas de piedemonte y zonas de inundación de las numerosas corrientes que drenan hacia el río Orinoco en dirección suroriente. De la vasta red de drenajes que se originan en el flanco oriental de la Cordillera Oriental colombiana, son de resaltar como principales a los ríos Arauca, Casanare, Meta y Guaviare, junto con otros de gran importancia como el Tomo, Bita, Vichada, Guainía, Tuparro, Inírida y Cinaruco (Cárdenas-Torres, 2011). Por otra parte, el piedemonte llanero es un área estratégica en cuanto a diversidad, por ser un punto de transición entre ecosistemas montanos y tropicales (pasando desde los bosques muy húmedos hasta los secos), haciendo de esta una zona de convergencia de diferentes ecoregiones (Andina, Amazónica, Orinoquía) (Carvajal et al., 2007).
La presente investigación se circunscribe al estudio de dos coberturas boscosas representativas de esta región: los bosques de galería (Bg) y los bosques densos altos de tierra firme (Bda). Los primeros son considerados ecosistemas de importancia para la conservación del recurso hídrico en áreas de planicie asociados a cursos de agua, con altos niveles freáticos (inundables en invierno) que actúan como corredores de dispersión y albergue de la fauna silvestre; adicionalmente, estos ecosistemas proveen el hábitat a invertebrados que son fuente importante de alimento para la fauna acuática y terrestre (Valencia, 1993). Los bosques densos altos de tierra firme configuran otro tipo de ecosistema con factores y recursos distintos, nivel freático bajo (no inundables todo el año) y en la mayoría de los casos constituyen fuentes o nacimientos de agua, los cuales se ciñen a pequeños parches aislados o “islas” en predios privados, siendo en muchos casos el producto de cultivos abandonados de cacao (Theobroma cacao L.) o de café (Coffea arabica L.), donde se permitió que las dinámicas de sucesión natural se encargaran de establecer los actuales bosques. Se presume que ambos cuentan con composición florística, estructura y diversidad distinta, pudiendo albergar organismos que, en algunos casos, se encuentran catalogados en categorías de amenaza de extinción.
En este sentido, resulta de consideración que los procesos de desarrollo socioeconómico en la región de los Llanos Orientales han desencadenado un detrimento sustancial en la existencia de la masa arbórea natural en ambos ecosistemas, llevándolos a importantes niveles de fragmentación que ponen en riesgo la estabilidad de otros recursos asociados, como la calidad y cantidad de agua, la afectación sobre el hábitat de especies de fauna silvestre (en algunos casos en amenaza de extinción) y, en general, la disminución en la oferta de bienes y servicios, poniendo en peligro, al mismo tiempo, elementos de la flora local. Adicionalmente, y para el caso de los bosques de galería, los servicios ecológicos que prestan estas comunidades son considerados de gran importancia, pues una comunidad conservada sirve de filtro entre el río y los ambientes adyacentes, impide el flujo al torrente del río de agroquímicos y productos orgánicos utilizados como insumos agrícolas y desechos agropecuarios, además de amortiguar algunos de los procesos de sedimentación de los lechos de los ríos.
Dichos servicios ecológicos mantienen la calidad del agua y proveen protección contra las inundaciones y la erosión (Timoney et al., 1997). Por ello resulta fundamental el conocimiento de su estado actual, considerando el alto deterioro evidenciado en estos ecosistemas que aún siguen cumpliendo con importantes funciones ambientales, pero especialmente para valorarlos como espacios naturales que aún albergan objetos de conservación.
El objetivo del trabajo es comparar la composición florística, estructura y diversidad de especies forestales halladas en ecosistemas de bosque de galería y bosque denso alto de tierra firme en un sector de los municipios de Acacías y Guamal (Llanos del Orinoco colombiano) y determinar la presencia de especies forestales amenazadas.
MATERIALES Y MÉTODOS
LOCALIZACIÓN
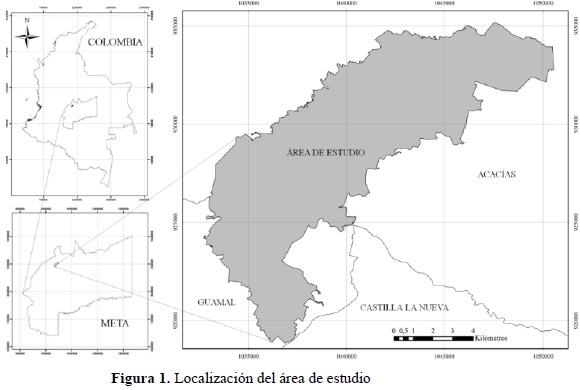
El área de estudio se encuentra localizada en la región geográfica de los Llanos Orientales colombianos, específicamente en los municipios de Acacías y Guamal del departamento del Meta, con influencia directa de la zona de piedemonte asociada a la vertiente oriental de la Cordillera Oriental (Figura 1). La región donde fue desarrollada la investigación se caracteriza por encontrarse en la zona de vida de bosque húmedo tropical propuesta por Holdridge (1978) a 500 m de altitud, donde predomina una temperatura promedio de 23°C y precipitaciones que se encuentran por el orden de los 2 700 mm anuales.
MAPA DE COBERTURAS VEGETALES BOSCOSAS
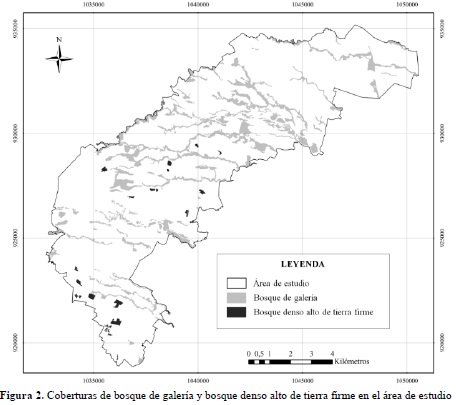
Durante la primera fase (en oficina), se elaboró un mapa de coberturas vegetales boscosas a escala 1:10 000 del área de estudio, que comprende 10 465 ha, donde se identificaron y mapearon las unidades cartográficas de bosque de galería y bosque denso alto de tierra firme con 1 229.94 y 60.45 hectáreas respectivamente; como resultado se obtuvo un mapa temático tal como se ilustra en la Figura 2.
DISEñO DE LA INVESTIGACIÓN
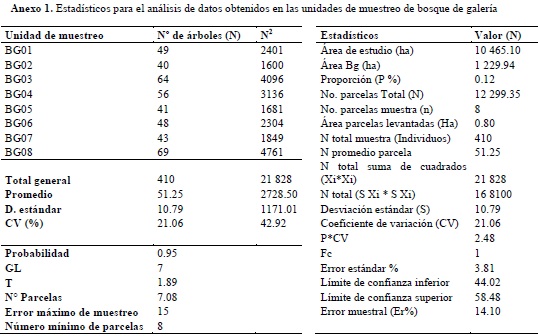
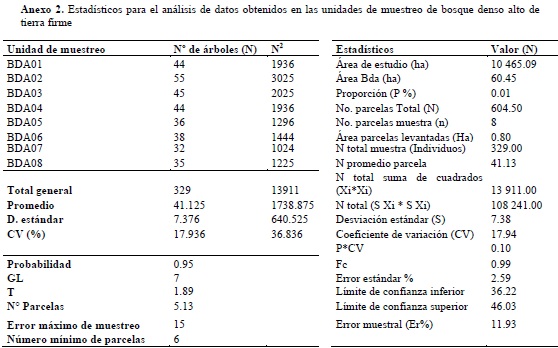
Se empleó un muestreo aleatorio estratificado (Melo & Vargas, 2003), donde se dividió la población en dos estratos (los dos ecosistemas en estudio), en los cuales se tomaron muestras al azar. Para ello fue empleado el mapa de coberturas donde se seleccionaron los sitios potenciales para la realización de los pre-muestreos mediante la elaboración cartográfica de una malla de puntos para cada unidad de análisis. Posteriormente, se hizo la selección, completamente al azar, de los puntos para el levantamiento de las parcelas en la fase de campo. Para realizar la comparación entre unidades de muestreo y comprobar que la información es representativa de la muestra, se tomó como variable el número de individuos, con el fin de procesar la información a una probabilidad del 95% y error de muestreo inferior al 15%. Los resultados del análisis de datos para calcular el tamaño de la muestra en bosque de galería y bosque denso alto de tierra firme, se encuentran consignados en los anexos 1 y 2, respectivamente.
TRAZADO Y MUESTREO DE LAS PARCELAS
Durante los muestreos forestales se tomaron datos en 8 parcelas de 0.1 ha (20 m x 50 m), por cada tipo de ecosistema, para caracterizar aquellos individuos con diámetros a la altura del pecho (DAP a 1.3 m del suelo) mayor o igual a 10 cm (censo de fustales), registrando datos de especie, altura total (HT) y altura a la base de la copa (HC), alturas que fueron medidas con el uso de clinómetro de suunto. De acuerdo con Cárdenas et al. (1997), este tipo de parcelas facilita un mayor número de muestreos en poco tiempo, permitiendo obtener una muestra representativa.
En cada punto localizado con GPS, se ubicó el norte magnético mediante el uso de brújula profesional y a partir de allí se extendió una cuerda por distancia de 50 metros y con marcas cada 10 metros. Esta línea constituye el eje de la parcela y a partir de esta se midieron todos los fustales que se encontraron 10 metros a ambos lados de la misma. Cada árbol fue marcado con un número consecutivo en pintura roja sobre una superficie visible y de fácil detección.
MEDICIÓN DE VARIABLES EN CAMPO
En los formularios se incluyen hasta 5 circunferencias a la altura del pecho (CAP), en razón a que algunas especies pueden presentar bifurcaciones por debajo de los 1.3 m del suelo y, consecuentemente, es necesario medir todas las CAP para estimar con precisión la CAP de cada individuo, que es el resultado de la ecuación a continuación descrita, empleada para casos de diámetros múltiples y propuesta por Ramírez & Kleinn (2001).
![]()
El DAP es empleado para establecer la dominancia e índice de Valor de Importancia (IVI) por especie, dentro del análisis de estructura horizontal; su cálculo se realizó con base en el CAP:
![]()
COLECTA, EMBALAJE Y PRENSADO DE MUESTRAS BOTÁNICAS
En el levantamiento de la información de campo se tomaron muestras botánicas para su posterior determinación en herbario, siguiendo la metodología propuesta por el Instituto de Investigación de Recursos Biológicos Alexander von Humboldt, en el Manual de métodos para el desarrollo de inventarios de biodiversidad (Villareal et al., 2006). La determinación taxonómica de los ejemplares botánicos se realizó en el Herbario Forestal de la Universidad Distrital de Bogotá (UDBC), Colombia. Los nombres científicos de las especies, así como su clasificación taxonómica, se corroboraron con base en las siguientes referencias electrónicas: The Plant List, The International Plants Name Index e Integrated Taxonomic Information System, las cuales fueron consultadas por última vez el 21 de febrero de 2014.
ESTRUCTURA HORIZONTAL
Consiste en determinar la frecuencia, área basal o dominancia, abundancia, e IVI de los individuos en la superficie del suelo (Becerra, 1971). Para el cálculo de estas variables se emplearon las fórmulas que se presentan en la tabla 1.

ESTRUCTURA VERTICAL
Se empleó el método cuantitativo de descripción de la vegetación propuesto por Ogawa, confeccionando una gráfica con las alturas totales en las ordenadas y las alturas a la base de la copa en las abscisas; la aparición de enjambres de puntos más o menos aislados, indica el virtual vacío de las copas en los niveles intermedios, sugiriendo un número de estratos diferenciales en el perfil del bosque. Cuando se genera una sola nube de puntos alargada y con pendiente positiva, no se pueden diferenciar estratos, ya que existe una continuidad de puntos desde el sotobosque hasta el dosel (Unesco, 1980).
Para la elaboración de los perfiles de vegetación fue necesario seleccionar dos parcelas tipo por cada ecosistema y luego se diagramaron las características principales de cada una de las especies halladas en estas y, posteriormente, con los puntos GPS de cada individuo se ubicaron los árboles en su correspondiente lugar en el espacio.
DIVERSIDAD
La información sobre el número de especies y de individuos presentes en cada ecosistema se empleó para calcular el cociente de mezcla, el índice estructural de diversidad de Shannon-Wiener, el índice de equidad de Simpson y el índice de riqueza de especies de Margalef.
ESPECIES AMENAZADAS, CASI AMENAZADAS Y CON PREOCUPACIÓN MENOR
Para la verificación de estas especies se consultó la Lista de Especies en Libros Rojos de Colombia, en la Resolución 0192 de 2014 del Ministerio de Ambiente y Desarrollo Sostenible (MADS), en la RedList de la UICN y la base de datos CITES.
ANáLISIS ESTADÍSTICOS
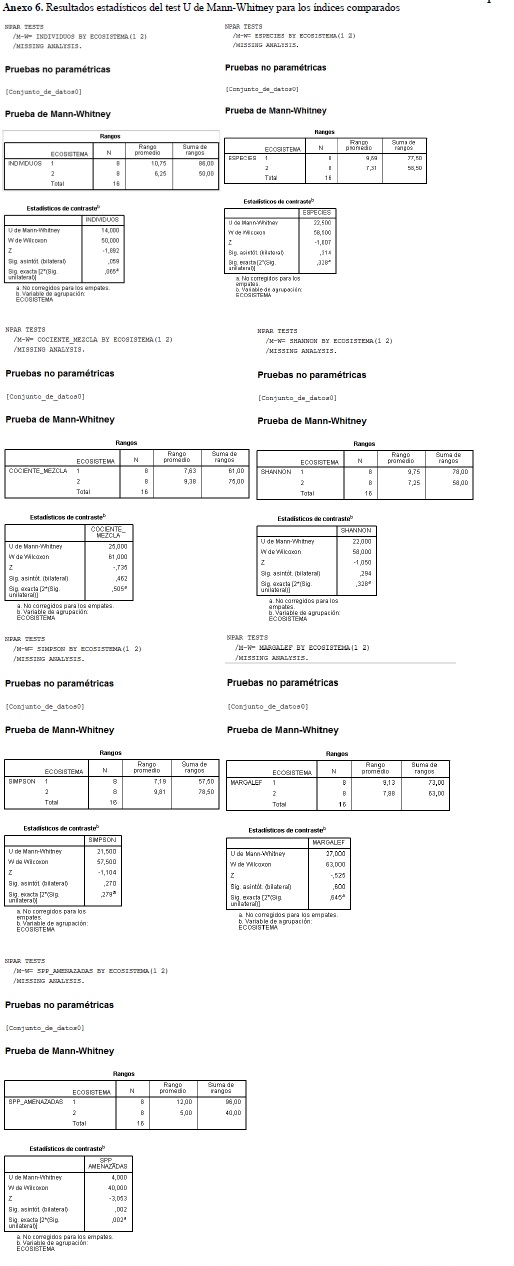
Dado el pequeño tamaño de la muestra (8 parcelas por ecosistema) y la distribución no normal de las variables analizadas, se excluyó la posibilidad de transformar los datos, que fueron analizados mediante el test no paramétrico U de Mann-Whitney para muestras independientes (Zar, 1984). La prueba se utilizó para comparar las siguientes variables entre ecosistemas: riqueza específica, abundancia absoluta, cociente de mezcla, los distintos índices de diversidad calculados y la cantidad de especies amenazadas, casi amenazadas y en preocupación menor. Para ello se empleó el programa IBM SPSS Statistics 19.
RESULTADOS
Con base en la información estandarizada para cada tipo de bosque, se realizaron los análisis comparativos de: composición florística, estructura horizontal (que incluye abundancia, frecuencia, dominancia e índice de valor de importancia - IVI), estructura vertical, donde se analiza el diagrama de Ogawa y los perfiles de vegetación, así como las características de diversidad evaluadas mediante los índices de Shannon-Wiener, Simpson y Margalef. Los análisis incluyen la determinación de la presencia-ausencia de las especies que se encuentren con algún grado de amenaza de acuerdo con las listas rojas de especies amenazadas a nivel nacional y global.
COMPOSICIÓN FLORÍSTICA
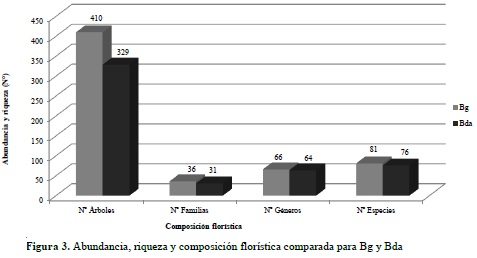
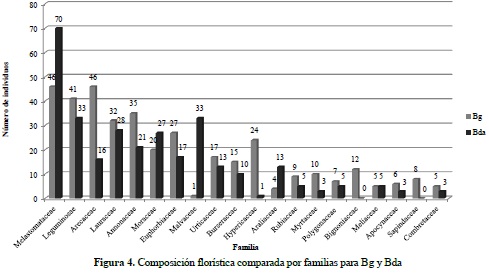
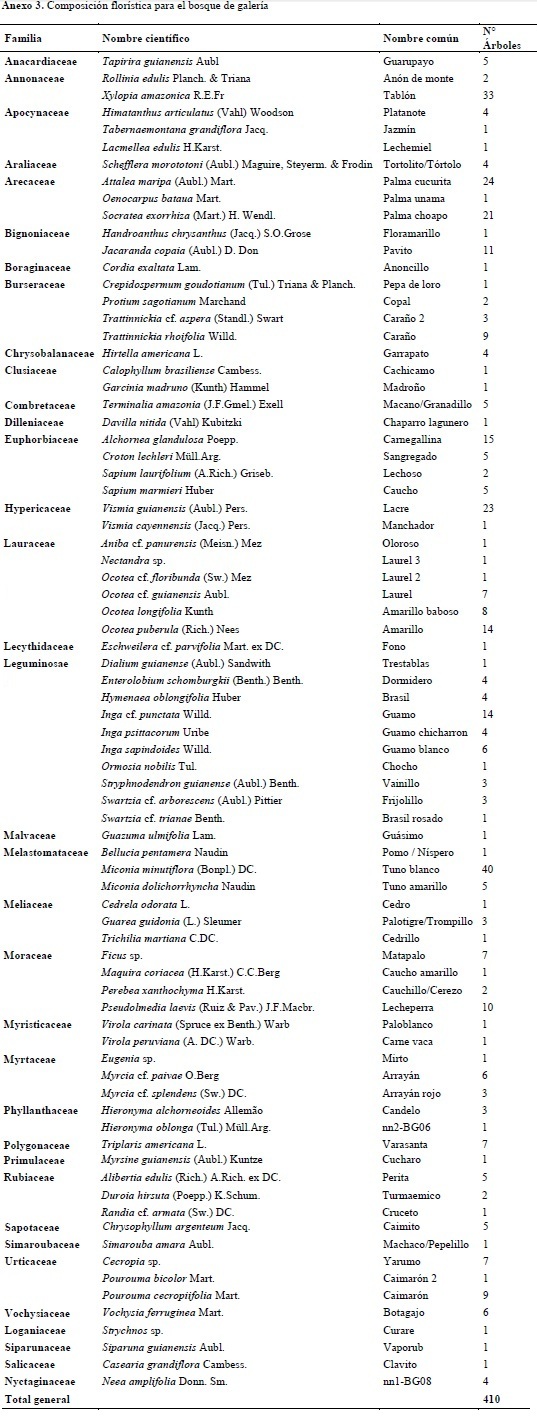
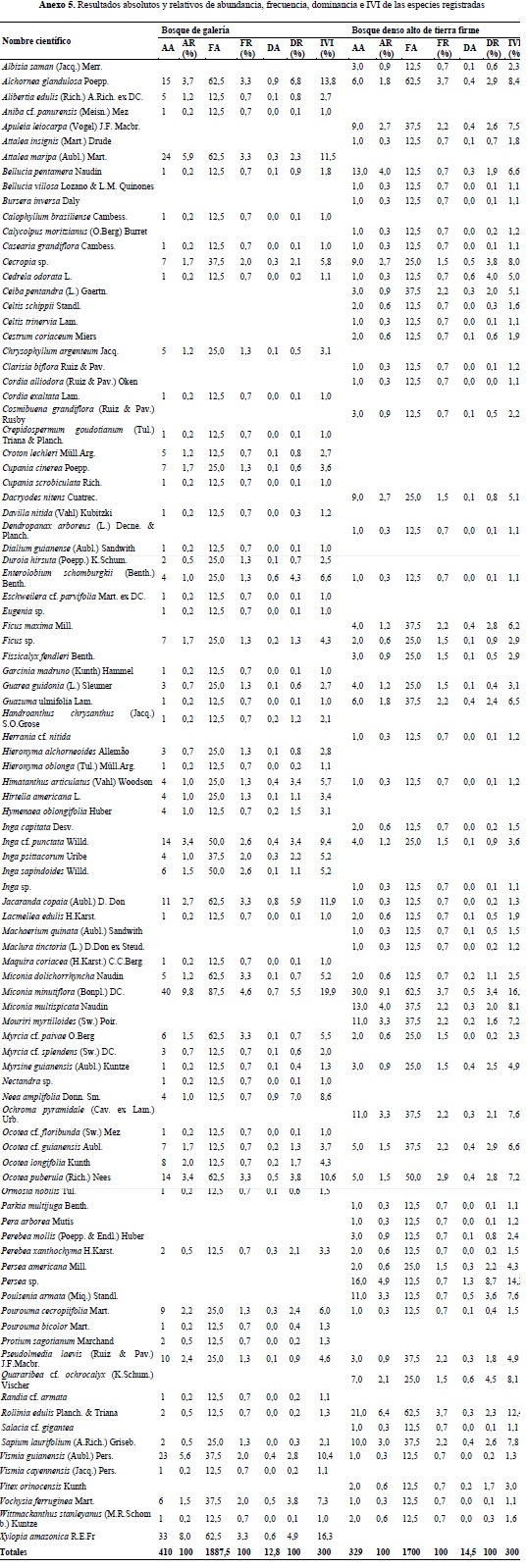
Este componente hace referencia al análisis de los distintos grupos de fustales presentes en las unidades de muestreo realizadas para cada tipo de bosque. Para el bosque de galería se encontró un total de 410 individuos y 81 especies, distribuidas en 36 familias y 66 géneros (Anexo 3); para el bosque denso alto de tierra firme se encontraron 329 fustales de 76 especies, 31 familias y 64 géneros (Anexo 4). En la Figura 3 se compara la abundancia, riqueza y composición florística detectada en los ecosistemas objeto de estudio, mientras que en la Figura 4 se compara la composición con base en las 15 familias que reportaron mayor abundancia de especies para ambos ecosistemas.
ESTRUCTURA HORIZONTAL
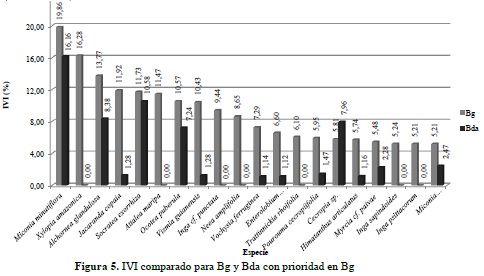
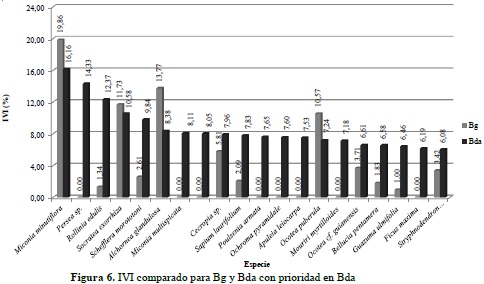
Para el análisis de la estructura horizontal se calculó el IVI en los dos ecosistemas, con base en la abundancia, frecuencia y dominancia de cada especie (anexo 5) y posteriormente fue graficado con las 20 más representativas, clasificadas en orden jerárquico para el Bg (Figura 5) y para el Bda (Figura 6). Los resultados permiten establecer una importante presencia de la especie Miconia minutiflora (Tuno blanco) para los dos tipos de bosque objeto de análisis. No obstante, Xylopia amazonica (Tablón) registra el segundo mayor IVI en Bg y no aparece dentro de las 20 más importantes en Bda; situación similar sucede con Persea sp. (Aguacatillo), segunda más importante en Bda y que tampoco se encuentra dentro de las más relevantes para Bg.
ESTRUCTURA VERTICAL
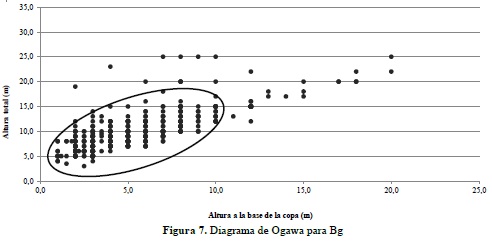
El diagrama de Ogawa permite establecer que la mayoría de los individuos registrados para el Bg se concentran en las clases altimétricas medias y bajas, con algunos individuos dominando los estratos superiores (Figura 7).
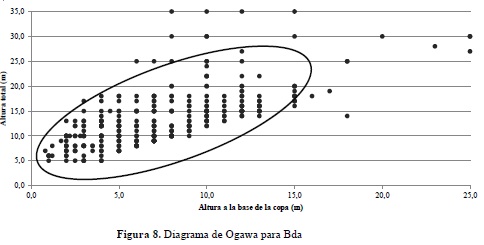
Para el caso del bosque denso, el diagrama de Ogawa arroja como resultado una vegetación mucho más homogénea en términos de distribución de las clases altimétricas (Figura 8).
Los ejemplos esquemáticos de las especies registradas en las parcelas de Bg y Bda se presentan en las figuras 9 y 10 a manera de leyenda y los correspondientes perfiles de vegetación se ilustran en las figuras 11 y 12. El perfil de vegetación elaborado para el bosque de galería permite establecer un estrato inferior caracterizado por la presencia de individuos con alturas menores a los 8 metros, donde destacan especies como Eugenia sp. (Mirto), Siparuna guianensis (Vaporub), Swartzia cf. arborescens (Frijolillo), Tabernaemontana grandiflora (Jazmín) y M. minutiflora. El estrato medio cuenta con la presencia de individuos entre los 8 y 15 metros de altura, agrupando una menor cantidad de estos comparado con el estrato inferior; en este estrato la especie M. minutiflora continúa siendo una de las más características, junto con T. americana, X. amazonica y algunas especies del género Ocotea. En el estrato superior se aprecia un dosel irregular con individuos de 25 m de altura de las especies T. americana, E. schomburgkii y algunas palmas como A. maripa y S. exorrhiza.
El perfil de vegetación del bosque denso alto de tierra firme permite diferenciar una significativa abundancia de elementos que se mantienen en una altura promedio de 15 metros, donde se destacan especies como Guazuma ulmifolia (Guásimo), Sapium marmieri (Caucho) y M. minutiflora; en el estrato inferior se encuentran especialmente individuos de T. cacao, lo cual confirma la información suministrada por los pobladores locales, quienes señalan que varios de estos bosques se originaron a partir de cultivos de cacao abandonados, suposición completamente válida si se considera que estos pueden coadyuvar en el crecimiento de la vegetación nativa al facilitar el desarrollo de las dinámicas de la sucesión natural temprana; dicha especie se encuentra acompañada por individuos de Alchornea glandulosa (Carnegallina), G. ulmifolia y otras del género Miconia. El estrato superior está dominado por las especies O. puberula, A. leiocarpa y especialmente por un individuo de C. odorata con más de 30 metros de altura.
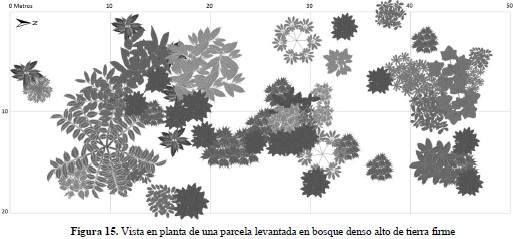
En el caso del Bg se puede apreciar que gran parte de los individuos se encuentran en alturas menores a los 10 metros; sin embargo, es posible diferenciar tres estratos arbóreos en estos bosques, donde predominan las especies M. minutiflora y Triplaris americana (Varasanta). En Bda se observa una estructura vertical mucho más alta y homogénea que la encontrada en el Bg, con elementos emergentes de gran porte representados por Ocotea puberula (Amarillo), Apuleia leiocarpa (Guacamayo) y Cedrela odorata (Cedro). Con el fin de facilitar la interpretación de los perfiles y la distribución de las especies en el espacio al interior de las parcelas seleccionadas en cada tipo de ecosistema, se presentan las vistas en planta (Figuras 13 y 15) con sus correspondientes leyendas que relacionan las especies con las formas de sus copas (Figuras 14 y 16).






DIVERSIDAD
El análisis comparado de la diversidad florística permitió establecer que el bosque de galería reporta el mayor número de especies (81) o riqueza específica (S) en el estado de crecimiento de fustal; igualmente, este registra la mayor abundancia absoluta (410). Con los anteriores datos se obtuvo un cociente de mezcla para Bg de 19.75%, revelando una menor heterogeneidad a la encontrada para el Bda que, con 76 especies distribuidas en 329 individuos, evidenció un cociente de mezcla de 23.10% (Tabla 2).
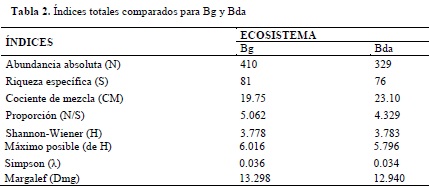
La proporción indica la relación entre la abundancia absoluta y la riqueza específica, señalando que en Bg es posible encontrar una especie por cada 5 individuos (1:5), mientras que en Bda se encuentra una especie por cada cuatro (1:4), confirmando de esta manera una mayor heterogeneidad para este último.
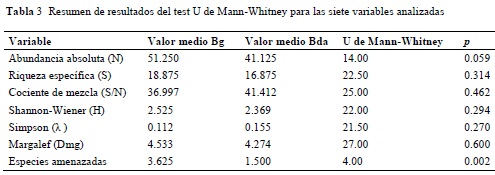
Los resultados en la aplicación de los índices de diversidad (para la totalidad de las unidades de muestreo) de Shannon-Wiener (3.778 y 3.783 respectivamente para Bg y Bda), de Simpson (0.036 y 0.034) y de Margalef (13.298 y 12.940), demuestran una gran similitud en la diversidad florística para los dos tipos de bosque. Sin embargo, no se encontraron diferencias significativas en la riqueza media de especies, ni tampoco en los índices de diversidad medios (Tabla 3 y
Anexo 6). Por otra parte, la abundancia absoluta tiende a ser distinta en ambos tipos de bosque, con una probabilidad levemente mayor a la esperada según el test (U = 14; P = 0.059).
ESPECIES AMENAZADAS, CASI AMENAZADAS Y CON PREOCUPACIÓN MENOR
Teniendo como fundamento la base de datos de las especies registradas dentro de las unidades de muestreo en los ecosistemas de Bg y Bda, se realizó un trabajo de revisión que permitió identificar su estatus o categoría de amenaza según la Lista de Especies en Libros Rojos de Colombia, la Resolución 0192 de 2014 del MADS, la base de datos de la RedList de la UICN y en la base de datos CITES (Tabla 4).
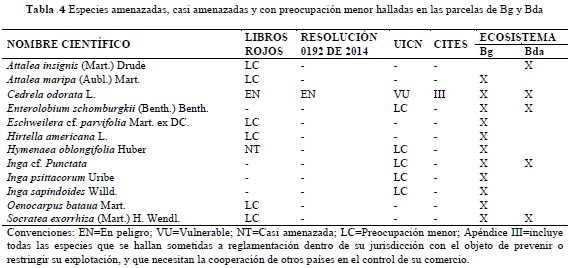
La aplicación del test estadístico reveló que existen diferencias significativas en el número de especies amenazadas, casi amenazadas y con preocupación menor entre ambos ecosistemas (U = 4; P = 0.002), con una probabilidad del 95% y con mayor significancia para el bosque de galería (Tabla 3).
DISCUSIÓN
La comparación del número de individuos por familia taxonómica permite establecer una clara dominancia de la familia Melastomataceae en ambos ecosistemas, con un total general de 116 registros, siendo más abundantes en el ecosistema de Bda (70) y en segundo lugar de importancia se destaca la familia Leguminosae con una riqueza específica de 74 individuos, de los cuales 41 se reportaron en Bg y 33 en Bda. Estas familias concentran el 25.71% del total de individuos inventariados, lo cual concuerda con las investigaciones realizadas por Carvajal & Murillo (2007), en su análisis florístico y fitogeográfico del sector nororiental de la Sierra de La Macarena, donde reportan a la familia Melastomataceae como una de las más diversas y abundantes en bosques de galería y bosques de tierra firme poco intervenidos, al igual que lo planteado por Duque et al. (2003).
El mayor IVI alcanzado por la especie M. minutiflora en los dos ecosistemas, coincide con los planteamientos anteriormente expuestos por los citados autores, demostrando una vez más la importancia de la familia Melastomataceae como una de las más diversificadas para este sector del piedemonte llanero.
En términos generales, y a pesar de no encontrar diferencias significativas en la biocenosis de estos dos ecosistemas, el análisis de la estructura horizontal permite establecer una clara divergencia entre el peso ecológico de las especies halladas en los inventarios; también se evidencia en la presencia-ausencia de algunas especies en un ecosistema y en otro, situación que puede darse principalmente por las características y disponibilidad de factores y recursos para el desarrollo de la vegetación en cada unidad de análisis, e indiscutiblemente por la intervención directa del hombre para el desarrollo de sus actividades productivas; es el caso de especies con una importancia ecológica considerable en Bg como lo son X. amazonica, A. maripa y Neea amplifolia, pero que no aparecen dentro de las más relevantes en Bda, mientras que Persea sp., Miconia multispicata y Quararibea cf. ochrocalyx, alcanzan un IVI por encima del 8% en Bda y no aparecen dentro de las más importantes en Bg. Este hecho puede estar asociado a las dinámicas de disponibilidad del recurso hídrico en ambos ecosistemas, con mucha mayor abundancia en bosque de galería, donde, por ejemplo, A. maripa se posiciona como una especie tolerante a periodos de inundación y muy abundante en bosques de ribera, según lo planteado por Díaz et al. (2010), en su estudio sobre composición florística del bosque ribereño del río San José, Reserva Forestal de Imataca, Estado Bolívar, Venezuela, y confirmado los resultados de Mostacedo et al. (2006), quienes establecen que A. maripa se encuentra dentro del 25% de las especies arbóreas raras en bosque alto.
La dispersión de las clases de altura en Bg revela que la mayoría de los individuos inventariados en las unidades de muestreo se encuentran acumulados en los estratos medio e inferior, es decir por debajo de los 10 metros, indicando cierto grado de alteración de estos ecosistemas, especialmente porque aparecen pocos individuos en el estrato superior alcanzando más de 20 metros de altura de algunas especies que no son apetecidas para su aprovechamiento forestal como es el caso de T. americana, cuyo uso principal es medicinal, así como las palmas A. maripa y S. exorrhiza, igualmente abundantes en el bosque de galería y que no demuestran un uso intensivo por parte de la comunidad. Por el contrario, el ecosistema de Bda presenta una distribución mucho más homogénea de los diferentes individuos en las clases de altura, lo cual presume una mayor estabilidad en las dinámicas sucesionales en estos bosques y teniendo en cuenta que se mantienen protegidos por iniciativas privadas con fines contemplativos y como reservas para el aprovechamiento fortuito de madera.
En los índices de diversidad calculados para los dos ecosistemas se detectó una amplia similitud entre estos. Patil & Taille (1982) mencionan que el índice de Simpson tiene la tendencia de ser más pequeño cuando la comunidad es más diversa; los valores calculados para bosque de galería y bosque denso (0.034 y 0.036 respectivamente) indican una muy baja probabilidad de que dos individuos tomados al azar sean de la misma especie (Melo y Vargas, 2003), lo cual significa que existe una alta heterogeneidad específica en estos dos ecosistemas y consecuentemente una alta diversidad florística; sin embargo, según el cociente de mezcla calculado, resulta interesante encontrar que el Bda cuenta con un mayor valor de heterogeneidad respecto al Bg.
La alta heterogeneidad florística es también apoyada por los valores del índice de Shannon-Wiener (3.778 y 3.783 para Bg y Bda) con relación a sus máximos posibles (logaritmo natural del número de individuos, con valores de 6.016 y 5.796 respectivamente (Margalef, 1995).
También los resultados del índice de Margalef apoyan esta interpretación. Se asume que valores de este índice, inferiores a 2, determinan un baja diversidad y superiores a 5 indican una alta diversidad florística (Margalef, 1995); para los fustales del Bg se encontró un valor de 13.298, lo cual representa un bosque bastante heterogéneo; similar resultado arrojó el cálculo para el Bda (12.940), a pesar de contar con una menor riqueza de especies, pero con una menor cantidad de individuos y mucho menos representatividad en superficie respecto al área de estudio, condición que lo posiciona como un ecosistema de importancia en términos de biodiversidad.
De esta manera, los índices de diversidad calculados para estos ecosistemas permiten establecer que ambos poseen una gran similitud en cuanto al número de individuos versus el número de especies, muy posiblemente porque se trata de bosques circunscritos en la misma área geográfica del piedemonte metense, los cuales comparten las mismas condiciones de clima y pendiente, interactuando con elementos faunísticos de distribución compartida que necesariamente intervienen en las dinámicas de la sucesión natural.
De acuerdo con las bases de datos consultadas para establecer la presencia de especies catalogadas en alguna categoría de amenaza, las casi amenazadas y en preocupación menor, fue posible encontrar un total de 12, de las cuales 11 están registradas en Bg y 5 en Bda (Tabla 4); dicha situación impulsa a considerar estos ecosistemas como áreas de interés para la conservación, ya que a pesar de encontrarse restringidos a los cursos de agua en el caso de los Bg, a pequeños relictos boscosos a manera de “islas” para el caso de Bda, y teniendo en cuenta su alto grado de fragmentación, en los dos se encontraron especies amenazadas. Por citar un ejemplo, en ambos ecosistemas se registraron individuos de C. odorata, especie catalogada En Peligro de extinción (EN) a nivel nacional según la Lista de Especies en Libros Rojos de Colombia, considerada como Vulnerable (VU) A1cd+2cd por la RedList de la UICN y Apéndice III en CITES.
La presencia de esta especie en cada tipo de bosque inventariado es muestra de la necesidad de establecer estrategias de investigación en diversidad forestal, no solo para la protección de los relictos de bosque, sino también para impulsar el desarrollo de estudios científicos sobre estos ecosistemas, permitiendo el aprovechamiento de su potencial como fuente de material genético para futuros viveros que se establezcan en la zona y que fomenten su reproducción mediante el enriquecimiento de otras coberturas naturales como la vegetación secundaria, la cual también juega un papel importante en las dinámicas sucesionales de los bosques naturales al implicar una superficie de considerable extensión en el área de estudio.
CONCLUSIONES
Desde el punto de vista de riqueza comparada, la composición florística de los bosques de galería reportó mayores valores en las tres categorías taxonómicas analizadas: especies, géneros y familias. Dichas diferencias pueden explicarse por la mayor densidad de los fustales que en bosques densos (un 25% más de individuos registrados). Sin embargo, no existen diferencias significativas entre los valores medios de riqueza específica y abundancia absoluta, ni tampoco entre los índices de diversidad calculados para ambos ecosistemas (Tabla 3).
Es probable que las actividades humanas de la región jueguen un papel importante en la abundancia de individuos en los ecosistemas de Bda, los cuales, en la mayoría de los casos, forman parte de iniciativas de conservación privada, pero siendo vistos como reservorios de madera para su aprovechamiento ocasional en las infraestructuras asociadas a los pastos manejados, donde resulta indispensable, por ejemplo, reemplazar postes para cercas y la construcción de establos; ello teniendo en cuenta que la actividad ganadera es una de las principales en esta región del país.
El análisis realizado para comprender la arquitectura de ambos ecosistemas en un plano vertical resultó ser una variable importante para la presente investigación, ya que permitió visualizar una clara diferencia en la distribución de los individuos en altura, donde el bosque denso revela una mejor representatividad del arbolado en todos los estratos y consecuentemente un comportamiento típico de bosques poco intervenidos, contrario a lo que sucede con el Bg, el cual acumula la mayor cantidad de árboles en el estrato medio, con algunos elementos emergentes en el dosel, señales propias de bosques intervenidos y sometidos a procesos de fragmentación. En este sentido, para estudios comparativos de bosques naturales, el análisis de la estructura vertical es una variable a considerar en el momento de establecer diferencias en el desarrollo y estado sucesional de los mismos.
Un resultado destacable es el número de especies amenazadas, casi amenazadas y en preocupación menor, significativamente mayor en el bosque de galería que en el bosque denso, situación que puede darse especialmente por el alto grado de fragmentación evidenciado en este último, pero también por el mayor flujo de especies que se presenta en el bosque de galería al encontrarse asociado a cursos de agua. A pesar de las diferencias encontradas en estos dos ecosistemas, el solo hecho de que alberguen especies amenazadas en su interior los posiciona como áreas de interés para la conservación de los recursos naturales a diferentes escalas, incluyendo el fomento de estrategias de gestión como por ejemplo las Reservas Naturales de la Sociedad Civil, compra de predios por parte de la empresa privada y los municipios, así como otros instrumentos exitosos de conservación local y regional. Dichas estrategias pueden realizarse con especial atención en los bosques de galería al encontrarse mucho más expuestos a intervenciones por parte de foráneos, pero prestando igualmente atención al manejo de los bosques densos, teniendo en cuenta su alto grado de aislamiento y falta de conectividad con otras coberturas boscosas.
Este estudio comparativo permite concluir la gran similitud que existe entre bosques de galería y bosques densos, ya que los segundos, en algunos casos, son el resultado de la fragmentación de los primeros, dejando como consecuencia parches de bosques aislados y sin una conectividad ecosistémica con las áreas que continúan manteniendo mayores regímenes de inundación, es decir con los actuales bosques de galería. Esta interpretación se encuentra sustentada, además, en los resultados obtenidos mediante el test U de Mann-Whitney, donde no se encontraron diferencias significativas en los índices analizados; no obstante, fue posible establecer que estos procesos de fragmentación y las características propias de los tipos de bosque analizados, especialmente las asociadas a los regímenes de inundación, influyen en la abundancia de algunas especies tolerantes a niveles freáticos altos como A. maripa en bosque de galería y sin presencia en los bosques densos.
AGRADECIMIENTOS
Esta investigación fue desarrollada con información proveniente de los inventarios forestales realizados para el Estudio de Impacto Ambiental (EIA) del Campo de Producción 50k CPO-09, financiado por Ecopetrol S.A. y ejecutado por el Consorcio Megaoil. El autor expresa sus agradecimientos a la Ingeniera Luisa Ortiz de Ecopetrol S.A., al Ingeniero Vicente Pico, Representante Legal de Megaoil y a la Ingeniera Loyda Hernández, Coordinadora del EIA, por permitir el empleo de los datos para este trabajo de investigación. Al Dr. Tomás Santos, profesor de la Universidad Complutense de Madrid, por la revisión de textos y de análisis estadísticos. Al Dr. Manuel Merchán Fornelino, profesor de la I.E. Universidad (Segovia, España) y Presidente de la Asociación Chelonia Internacional, y a la Dra. Ana Fidalgo de las Heras, profesora de la Universidad Autónoma de Madrid, por la comprobación de los resultados estadísticos. Al Ingeniero William Ariza, Director del Herbario Forestal Gilberto Emilio Mahecha de la Universidad Distrital de Bogotá (UDBC) por la determinación de las muestras botánicas.
REFERENCIAS BIBLIOGRÁFICAS
Andrade-C., M.G. (2011). Estado de conocimiento de la biodiversidad en Colombia y sus amenazas. Consideraciones para fortalecer la interacción ciencia-política. Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales, 35 (137), 491-507.
Becerra, J. (1971). Notas de Ecología Forestal. Bogotá: Universidad Distrital Francisco José de Caldas. 69 p.
Bernal, R., Celis, M. & Gradstein S.R. (2007). Plant diversity of Colombia catalogued. Taxon, 56 (1), 273.
Cárdenas, D., Giraldo-Cañas, G. & Arias C. (1997). Vegetación. Capítulo 5. En Instituto Geográfico Agustín Codazzi (IGAC). Zonificación Ambiental para el Plan Modelo Colombo-Brasilero (Eje Apaporis-Tabatinga: PAT) (pp. 183-228). Bogotá: IGAC.
Cárdenas-Torres, M.A. (2011). Hábitat y distribución. En M. Merchán (ed.): Historia natural y conservación del caimán llanero (Crocodylus intermedius) en Colombia (pp. 57-123). Madrid: Asociación Chelonia.
Carvajal, L., Patarroyo J., Puentes D. & Valero J. (2007). Caracterización florística y estructural del piedemonte llanero en el departamento del Meta. Bogotá: Corporación para el Desarrollo Sostenible del área de Manejo Especial La Macarena (Cormacarena), Universidad Distrital Francisco José de Caldas. 130 p.
Carvajal, L. & Murillo J. (2007). Análisis Florístico y Fitogeográfico del sector nororiental de la Sierra de la Macarena. Bogotá: Universidad Distrital Francisco José de Caldas. 214 p.
Díaz, W., Rueda, J., Acosta, O., Martínez, O. & Castellanos, H. (2010). Composición florística del bosque ribereño del río San José, Reserva Forestal de Imataca, Estado Bolívar, Venezuela. Acta Botánica Venezuelica, 33 (1), 1-21.
Duque, A.J., Cárdenas D. & Rodríguez N. (2003). Dominancia florística y variabilidad estructural en bosques de tierra firme en el noroccidente de la Amazonia Colombiana. Caldasia, 25, 139-152.
Holdridge, L.R. (1978). Ecología basada en zonas de vida. Instituto Interamericano de Cooperación para la Agricultura (IICA). San José: Servicio Editorial del IICA. 216 p.
IBM Corporation. (2013). SPSS Statistics 19. U.S.A. IBM Corporation. ISBN: 3-900051-07-0. Recuperado de http://www-01.ibm.com/software/co/analytics/spss/
Maldonado-Ocampo J.A., Vari R.P. & Usma J.S. (2008). Checklist of the Freshwater Fishes of Colombia. Biota Colombiana, 9 (2), 143-237.
Margalef, R. (1995). Ecología. Barcelona: Omega. 951 p.
Melo, O. & Vargas R. (2003). Evaluación ecológica y silvicultural de ecosistemas boscosos. Ibagué: Universidad del Tolima, CRQ, Carder, Corpocaldas, Cortolima. 238 p.
Mostacedo, B., Balcazar, J. & Montero, J.C. (2006). Tipos de bosque, diversidad y composición florística en la Amazonia sudoeste de Bolivia. Ecología en Bolivia, 41(2), 99-116.
Ministerio de Ambiente y Desarrollo Sostenible. (2014). Resolución 0192 de 2014. Bogotá: MADS. 36 p.
Patil, G.P. & Taile C. (1982). Diversity as a concept and its measurement. Journal of the American Statistical Association, 77, 548-567.
Ramírez, C. & Kleinn C. (2001). Inventario Forestal Global -GFS-. Estudio Piloto en Costa Rica. Manual de Campo. San José: Centro Agronómico Tropical de Investigación y Enseñanza (Catie), FAO, Ministerio del Ambiente y Energía (Minae). 36 p.
Remsen, J.V., Jr., Cadena C.D., Jaramillo A., Nores M., Pacheco J.F., Pérez-Emán J., Robbins M.B., Stiles F.G., Stotz D.F. & Zimmer K.J. (2012). A classification of the bird species of South America. American Ornithologists Union. Recuperado de http://www.museum.lsu.edu/~Remsen/SACCBaseline.html
Sarmiento, J. (2007). La Familia Orchidaceae en Colombia. Actual Biol, (Supl. 1), 84.
Timoney, K.P., G. Peterson. & R. Wein. (1997). Vegetation development of boreal riparian plant communities after flooding, fire and logging, Peace River, Canada. Forest Ecology and Management Elsevier Science Amsterdam, 93, 101-120.
Unesco. (1980). Descripción, funcionamiento y evolución de los ecosistemas forestales tropicales. En Unesco, Pnuma y FAO. Ecosistemas de los Bosques Tropicales (pp. 127-162). Paris: Unesco y Cifca.
Valencia, R. (1993). Arizona riparian inventory and mapping project. In: Riparian Management: Common Threads and Shared Interests. Barbara Tellman et al. Tech. Coord. USDA. Forest Service. Rocky Mountain Forest and Range Experiment Station. General Technical Report RM-GTR-226 Fort
Collins Co. EUA. 242 p.
Villareal H., álvarez M., Córdoba S., Escobar F., Fagua G., Gast H., Mendoza F., Ospina M. & Umaña A.M. (2006). Manual de métodos para el desarrollo de inventarios de biodiversidad. Programa de Inventarios de Biodiversidad. Bogotá: Instituto de Investigación de Recursos Biológicos Alexander von Humboldt. 236 p.
Zar, J.H. (1984). Biostatistical analysis. Second edition. New Jersey: Prentice-Hall. 718 p.
ANEXOS





Licencia
Colombia Forestal conserva los derechos patrimoniales (copyright) de las obras publicadas, y favorece y permite la reutilización de las mismas bajo la licencia Creative Commons Atribución-CompartirIgual 4.0 Internacional por lo cual se pueden copiar, usar, difundir, transmitir y exponer públicamente, siempre que:
Se reconozcan los créditos de la obra de la manera especificada por el autor o el licenciante (pero no de una manera que sugiera que tiene su apoyo o que apoyan el uso que hace de su obra).

